![]()
![]()
![]()
![]()
![]()
 МОРСКАЯ
ФАУНА
МОРСКАЯ
ФАУНА
![]()
![]()
![]()
![]()
Надо заметить, что в эоцене море как будто само «приглашало» млекопитающих заселить его воды. Теплые заливы Тетиса кишели рыбой, обещая сытную жизнь, а ниша крупных морских хищников оказалась относительно свободной. Хотя акулы и крокодилы продолжали чувствовать себя очень неплохо, многочисленные в мезозое водные пресмыкающиеся – плезиозавры, мозазавры и им подобные – теперь исчезли из морских просторов.
![]()

Амбулоцетус - Так палеонтологи представляют себе амбулоцетуса, одного из первых китов. Точнее – существо, представляющее собой «звено в эволюционной цепи», соединяющей наземных млекопитающих начала кайнозойской эры и современных китообразных: синих, серых и горбатых китов, кашалотов, нарвалов, касаток и дельфинов – всего примерно 40 родов и 80 видов животных, бороздящих просторы Мирового океана.

Следующие важное открытие было сделано в 1992 г. Команда американца
Ханса Тьюиссена (Hans Thweissen) открыла в Северном Пакистане, в слоях
возрастом около 48 млн лет, почти полный скелет «переходного звена»
между современными китами и их наземными предками. Большие веслообразные
ступни и мощный хвост животного свидетельствовали о том, что оно было
хорошим пловцом, а крепкие кости ног и подвижные локтевые и запястные
суставы – о том, что оно неплохо передвигалось и по суше. «Переходное
звено» получило название амбулоцетуса (Ambulocetus natans), «ходящего и
плавающего кита».
А еще позже
Тьюиссену, Джиндеришу и другим палеонтологам удалось найти окаменелости,
позволявшие восстановить последовательные стадии перехода китов от
околоводного к океаническому образу жизни. Были найдены остатки
длинномордых ремингтоноцетид (Remingtonocetids, например кутхицетуса –
Kutchicetus) и протоцетид (Protocetids, например родоцетуса – Rodhocetus),
также произошедших от каких-то «пакицетоподобных» предков на берегах
Тетиса. От протоцетид произошли уже вполне «дельфинообразные» дорудоны (Dorudon)
– возможные предки базилозавров и современных китов, постепенно
расселившихся по всем морям земного шара.
На
протяжении следующего за эоценом олигоценового периода уровень моря
понизился. «Прото-Индия» соединилась с Азией (результатом этого
«столкновения» стало возникновение Гималаев), а Австралия и Антарктида
удалились друг от друга, в результате чего образовалось широкое
свободное кольцо морей в Южном полушарии. Возникло южное циркумполярное
течение, начал образовываться ледяной панцирь. Это создавало новые
условия для живущих в морях млекопитающих, что, по мнению ряда
специалистов, и привело к возникновению современных подотрядов – усатых
и зубатых китов. Наиболее древней известной переходной формой между ними
и древними археоцетами является Llanocetus, первичный усатый кит,
найденный в антарктических отложениях возрастом около 34 млн лет. Судя
по всему, он вполне мог питаться крилем. Зубатые же киты, по мнению
специалистов, возникли примерно в то же время, развивая способности к
эхолокации, которая позволяла активно охотиться в глубине.
К сожалению,
находки остатков первых представителей двух современных отрядов крайне
редки. Понижение уровня моря в олигоцене осушило прибрежные районы,
которые могли содержать эти остатки, и они подверглись разрушению. Но
раскопки в более поздних слоях показывают, что немного времени спустя,
30 млн лет назад, настоящие усатые и зубатые киты были представлены
несколькими семействами.
Находки же более древних археоцетов и
их предков позволяли восстанавливать и пути становления адаптаций
китообразных к постоянной жизни к океане.
Как было сказано, уже
пакицетус имел определенные преобразования в слуховой системе,
характерные для китов. Тьюиссен предположил, что он мог пользоваться
ими, чтобы воспринимать звуки, распространяющиеся в грунте. Будучи
засадным хищником, поджидающим жертву в воде, он мог слышать шаги
возможной добычи, приближавшейся к водопою. Вполне возможно, также
поступал и амбулоцетус, в челюсти которого уже имелось начало
характерного для китов канала, проводящего звук к уху. Положив нижнюю
челюсть на грунт – как это делают крокодилы – амбулоцетус «лоцировал»
передвижение по берегу своих потенциальных жертв.
Луо (Zhe-Xi Luo) –
палеонтолог, сотрудник Музея естественной истории в Питтсбурге, показал,
что у базилозавров и дорудонтов – первых полностью водных китов –
слуховая система по строению была уже достаточно близка к слуховой
системе современных китов. Позже эволюция двух групп – зубатых и усатых
китов – пошла разными путями. Первые развили особенности, необходимые
для произведения и восприятия высокочастотных звуков, используемых при
эхолокации, а вторые «специализировались» на низкочастотных акустических
сигналах, позволявших общаться друг с другом на больших расстояниях.
Около 28 млн лет назад ранние зубатые киты уже имели некоторые из
костных структур, необходимых для восприятия высокочастотных звуков и,
соответственно, определенную способность к эхолокации.
Другие важные
изменения черепа в процессе эволюции китообразных включали перемещение
глазниц с верхнего (как у крокодилов) положения у пакицетуса и
амбулоцетуса на бока головы, как у протоцетид и современных китов.
Ноздри сместились с вершины морды у пакицетуса на вершину головы
(дыхало) у современных китов. Зубы стали простыми и однообразными –
приспособленными только для удержания, а не разжевывания добычи. Усатые
же киты, как известно, в процессе эволюции вообще утратили зубы; их
«китовый ус» – роговые пластины, с зубами никак не связаны.
Анализ
изотопного состава атомов кислорода, присутствующих в зубах ископаемых
китов, позволяет делать заключения о том, жили ли они в пресной или
морской воде – во второй присутствует большая доля изотопа 18О.
Получилось, что организм пакицетусов получал только пресную воду,
амбулоцетусы могли жить как в пресных, так и в соленых водоемах, а
протоцетиды были уже настоящими морскими животными.
Одна из наиболее важных адаптаций, характерных для китов, – их способность производить вертикальные удары хвостом, обеспечивающие быстрое плавание. Строение тазового пояса амбулоцетуса еще позволяло животному поддерживать свой вес при передвижении по суше, хотя непропорционально большие задние конечности этого животного и веслообразные ступни делали такое передвижение затруднительным. Но такие конечности очень неплохо подходили для гребли при передвижении в воде! У родоцетуса, представителя протоцетид, кости нижнего отдела передней конечности были сжаты и уже напоминали ласты, а длинные нежные ступни, возможно были перепончатыми. Связки между позвонками, формирующими крестец, у родоцетуса были ослаблены, что позволяло позвоночнику изгибаться так, чтобы создавать волнообразные вертикальные движения хвоста. По мнению Джинджериша, он плавал «по-собачьи» на поверхности, а под водой передвигался за счет совмещения толчков веслообразных задних ног и хвоста. Скорее всего, этот зверь еще не полностью порвал с наземной средой и периодически выходил на сушу, где передвигался толчками, – наподобие современных ушастых тюленей.

Родоцетус
Базилозавры и дорудонтиды были, как уже сказано, полностью водными созданиями. Как у современных китов, плечо у них оставалось подвижным, а локоть и запястье образовывали передний плавник. Задние ноги и тазовые кости были маленькими и слабыми. Однако вопрос о том, когда именно киты окончательно потеряли задние конечности, остается открытым. Например, у настоящего усатого кита, чьи остатки были совсем недавно обнаружены в слоях возрастом 27 млн лет, еще имелись неплохо сформированные ноги…
В хвостовом отделе дорудона имелся позвонок округлой формы, подобный тому, который имеется у современных китов в основании хвостового плавника. Так что, возможно, дорудоны и базилозавры уже имели вполне китовый хвостовой плавник.
Итак, серия палеонтологических открытий 70–90-х гг. XX в. позволила проследить основные этапы эволюционного становления китов на их пути от наземных к исключительно водным животным. Однако вопрос об их более далеких предках продолжал – да и продолжает – оставаться открытым. Сравнение состава различных участков ДНК современных млекопитающих – представителей разных отрядов – позволил исследователям вновь говорить о высокой степени родства китов и парнокопытных. В то время как мезонихиды с их неясным систематическим положением оказывались вроде бы и ни при чем… Другие анализы показали большое сходство китов с отдельной и весьма специфической, надо сказать, группой парнокопытных – бегемотами
В палеогеновых морях, появились и другие млекопитающие, которые заняли освободившиеся экологические ниши морских рептилий. Это были древнейшие китообразные (Cetacea), остатки которых обнаружены в среднеооценовых отложениях Северной Африки (т. е. также в области эпиконтинеитальных морей океана Тетис). Пропорции тела архаических китов (Archaeoceti) значительно отличались от таковых современных групп: туловище было длинное и относительно тонкое (у наиболее крупной формы зейглодона — Zeuglodon — оно достигало в длину до 21 м при диаметре не более 2,5 м).
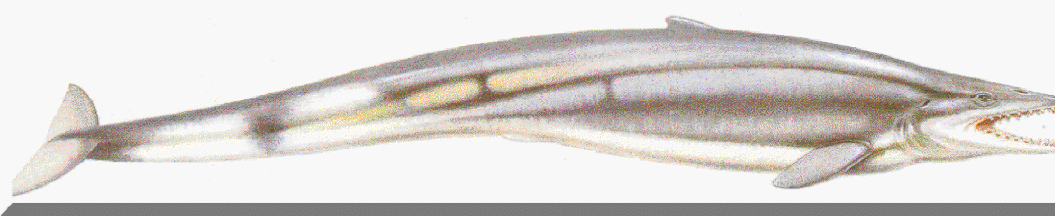
Длинные узкие челюсти большого (длиной до 1,2 м) и низкого черепа были усажены впереди коническими, а сзади сжатыми с боков острыми зубами. Короткие передние конечности располагались близко к голове, а задние были очень сильно редуцированы. По всей вероятности, китообразные возникли от каких-то примитивных наземных хищников — креодонтов, перешедших к рыбоядности и постепенно разливших приспособления к амфибиотической, а потом и к чисто водной жизни. Примитивные представители современных групп китообразных — зубатых (Odontoceti) и беззубых (MysLacoceti) китов — появились, соответственно, в позднем эоцене и олигоцене (Яковлева, 1993).

Первые ископаемые киты известны со времен эоцена. По всей видимости, они
пытались таким образом избежать обострившейся конкуренции на суше.
Ближе к концу эоценовой эпохи в океанах обитали существа, уже очень похожие на современных китов. Киты прекрасно адаптировались к жизни в воде. Обтекаемое туловище, широкие передние плавники и мощные горизонтальные хвостовые плавники делают их превосходными пловцами. Самые быстрые киты способны развивать скорость до 60 км/ч. Эти животные никогда не выходят на сушу, даже спариваются и рожают только в море.
Вода примерно в 800 раз плотнее воздуха. Она оказывает мощное лобовое
сопротивление любому животному, пытающемуся в ней передвигаться. Чтобы
уменьшить это сопротивление, морские животные должны иметь обтекаемую
форму тела. Организм кита вырабатывает особое жи ровое
вещество, ворвань, которое придает его телу гладкость, заполняя любые
полости под кожей
и сглаживая любые выпуклости и неровности на ней. Еще большая
обтекаемость достигается
за счет уменьшения выпирающих частей тела. Задние конечности китов
полностью исчезли, наружных ушных раковин у них тоже нет. Пенис самца
полностью скрыт в мышечных складках, а соски самки убраны в специальные
"карманы". Все это обеспечивает китам идеальную обтекаемость. Есть и еще
приспособление, помогающее китам так быстро плавать. Недавно было
установлено, что они ускоряются, как бы выпрыгивая из собственной кожи.
Двигаясь вперед, киты оставляют за собой что-то вроде "призрака" из
очень тонкого внешнего слоя кожи. Вода давит на этот "призрак", а само
животное проскакивает вперед — примерно так же, как кусок влажного мыла
выскальзывает из ваших мокрых рук, когда вы принимаете ванну.
ровое
вещество, ворвань, которое придает его телу гладкость, заполняя любые
полости под кожей
и сглаживая любые выпуклости и неровности на ней. Еще большая
обтекаемость достигается
за счет уменьшения выпирающих частей тела. Задние конечности китов
полностью исчезли, наружных ушных раковин у них тоже нет. Пенис самца
полностью скрыт в мышечных складках, а соски самки убраны в специальные
"карманы". Все это обеспечивает китам идеальную обтекаемость. Есть и еще
приспособление, помогающее китам так быстро плавать. Недавно было
установлено, что они ускоряются, как бы выпрыгивая из собственной кожи.
Двигаясь вперед, киты оставляют за собой что-то вроде "призрака" из
очень тонкого внешнего слоя кожи. Вода давит на этот "призрак", а само
животное проскакивает вперед — примерно так же, как кусок влажного мыла
выскальзывает из ваших мокрых рук, когда вы принимаете ванну.
Морская ихтиофауна палеогена уже включала все ныне существующие крупные группы рыб, хотя видовой состав и отличался от современного. Уже в плиоценовой эпохе неогена существовала крупнейшая из хищных акул Megalodon, длина тела которой достигала 30 м, а диаметр открытой пасти был свыше 2 м.
